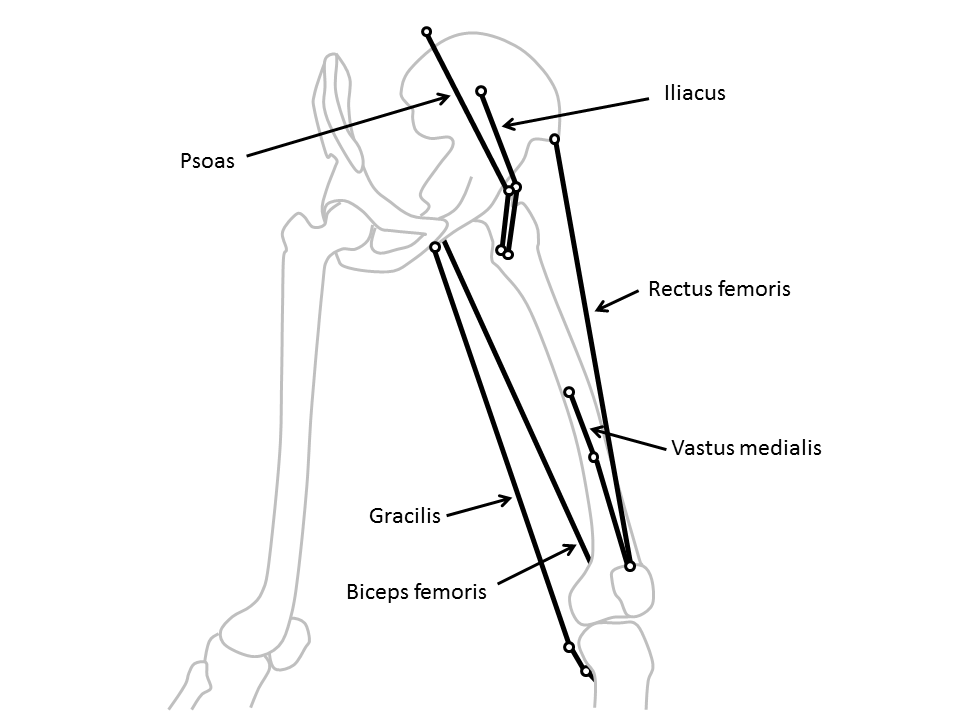
Lurking somewhere on this blog-site is software that Vicon users can download to calculate “muscle lengths”. It’s based on calculating the distance between the origin of a muscle on one segment and its insertion on another as illustrated in the diagram below (taken from my book). Some of the muscles (such as the rectus femoris) can be best represented as a straight line between the origin and insertion, whereas others (such as psoas or iliacus) have to pass around bones and may be better represented by including a “via point” along the path and adding the lengths of the two lines thus created.
Clearly as the joint or joints linking the relevant segments move this distance changes and you can thus plot muscle length on gait graphs in the same way that you can plot any other gait data. The technique has been around for a very long time but has been particularly popular since Scott Delp’s work on SIMM in the late 1980s.
Recently Jussi has added a comment to the page first saying that he’s got the software working (good) but then that a “somebody” has suggested “that ‘point-to-point’ muscle length models tend to be inaccurate, and a joint angle/moment arm based methods would be more accurate”.
This is quite an interesting comment because I don’t really consider this technique as “accurate”. The technique is based on a rather crude scaling of one set of origin and insertion coordinates. We don’t really know how consistent these are across healthy individuals and certainly not how they are affected in people with the sorts of conditions that we generally assess in clinical gait analysis (particularly those with bone, joint or muscle deformity). Further the calculations are dependent on the assumptions you make about how the joints move and ultimately on the accuracy of the joint angle measurements. All in all this is probably best described as a technique to “estimate” muscle length rather than to “calculate” it.
My general advice for clinical interpretation is that if you are dealing with single joint muscles then the muscle length graphs don’t really tell you much that you can’t already see on the joint angle graphs. Generally as a the joint extends the extensors get shorter and the flexors get longer (and vice versa) and the muscle length graph looks extremely similar to the corresponding joint angle graph (but with different units). Given that the actual calculations of muscle length are subject to so many assumptions you might as well work directly from the joint angle graphs.
The multi-joint muscles are different though because the muscle length depends on the orientation of both joints and the separate moment arms of the muscle about each. It is thus virtually impossible to assess how the muscle length varies through the gait cycle. In this case muscle length graphs are the only sensible way of getting an insight into how a muscle is behaving and can be valuable despite knowing that the actual values are only estimates. At least they are consistent estimates so that there is some sense in comparing the data you estimate for a patient against normative data which you have estimated using the same modelling procedure.
The most obvious example of this is in considering hamstrings length in children with cerebral palsy. It is extremely tempting to see a bent knee and assume that the hamstrings must be short. The “obvious” surgical response is to lengthen them. In many kids, however, the hip is also flexed and, because the moment arm at the hip is greater than that at the knee the muscles is often actually considerably longer than “normal“. This would suggest that surgical lengthening is inappropriate. Scott Delp and Alison Arnold drew the attention clinical community to this nearly twenty years ago and if there is one good reason for including muscle length estimates in gait reports for kids with CP then this is it. The data doesn’t have to be that accurate to be a reminder to surgeons that this is an important issue.
In direct response to Jussi’s question I don’t think its possible to say whether “point-to-point” or moment arm based calculations are more accurate. The calculations are affected by different factors and its not at all clear whether either is superior in general. The accuracy of whichever technique you use will be dependent on the quality of the input data (point coordinates or moment arms). As pointed out above accuracy is limited by a number of other factors and some of these may be more important than the choice of technique. Perhaps most importantly I’m not aware of any research that has ever been done to assess the accuracy of any muscle length calculations (though there is at least one that investigates the difference in results from a using a number of techniques ).
Of course this assumes that muscle lengths are being used clinically to understand why a patient walks the way they do. Anyone wanting to use them for more technical purposes perhaps in the generation of more advanced muscuol-skeletal models really needs to develop an in depth appreciation of all these factors for themselves.
.
PS Just to avoid the terminology police its worth reminding people that what almost everyone refers to as “muscle length” is actually the musculotendinous unit length. Maybe this is something that should have been added to my rant on terminology last week.