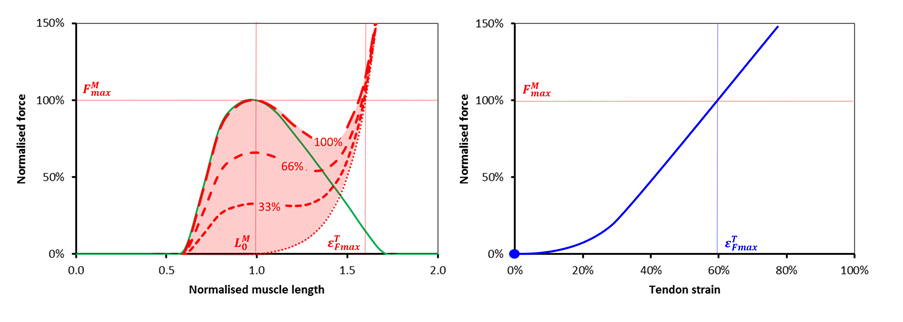
One of our students studying for the MSc in Clinical Gait Analysis recently wrote an essay of “The role of gait analysis in spina bifida” (myelomeningocele) which reminded me about work we did about 15 years ago when I was in Belfast (unfortunately it only ever got written up in Michael Eames’ MD thesis). At the time we were doing some modelling to look at how people with this condition walk. This young man has got a lesion at lower lumbar level (L4/L5) which means he’s got marked weakness in a range of lower limb muscles including plantarflexors, hip extensors and hip abductors. (He’s walking with the help of solid AFOs although you don’t see these on the “skeleton”). Importantly he retains function in his quadriceps (which are innervated from above this level).
This is a classic gait pattern for a person with this condition. It shows the trunk moving considerably from side to side. It looks clumsy and uncontrolled and its extremely tempting to make straightening him up an aim of therapy.
But the data were collected as part of a study into how the centre of mass moves. If you look carefully you’ll see there appears to be a blue ball moving around within the pelvis. This is the centre of mass, the point at which all the body’s weight can be assumed to act. It appears to be moving a lot from side to side confirming how much the trunk is moving around. If you hover your mouse pointer over it, however, you’ll find this is an optical illusion. The ball itself is moving very little from side to side, it only appears to be moving because of the larger movements of the trunk and especially the pelvis which your eye tends to follow. When we calculated the excursion from side to side it was very little more than the excursion that you or I use when walking around.
So what we might be seeing is not clumsy and uncontrolled gait but an exquisitely controlled gait that keeps his centre of mass pretty much in the same place whilst there is all that trunk movement going on around it. This is particularly amazing given that he has no functioning in his abductor muscles which are those that are primarily responsible for medio-lateral control. What we found is that his trunk moves so far that, at the end of the range of movement when control is required, the adductor muscles, which are preserved at this level, are able to provide this control. An alternative way of looking at this gait pattern is as the central nervous system is doing an amazing job in optimising the way he walks taking into account his neuromuscular deficits. If you try and train him to walk more upright the chances are he’ll walk a lot less efficiently and perhaps less functionally. I’m not even sure he could walk at all without the trunk sway (and without walking aids). If you want the optimum gait pattern given his impairments maybe you should leave the gait training to the central nervous system!
It’s for this reason that I prefer to talk about walking with pathology rather than pathological walking. I suspect that most of our patients are walking as optimally as they can given their impairments (which might included impairments to the central nervous system). Their walking is anything but pathological.
Of course the gait pattern may place abnormal stresses on various structures which may cause degenerative changes. This needs to be factored into any management plan but it should also be remembered that people with gait patterns like this quite often do not walk very much and the potential detrimental effects of high loading during walking may be offset to some extent by its limited duration.